Science 2 October 2009:
Vol. 326. no. 5949, pp. 64, 75-86 Science誌本文
DOI: 10.1126/science.1175802
Ardipithecus ramidus and the Paleobiology of Early Hominids
2 Rift Valley Research Service, Post Office Box 5717, Addis Ababa, Ethiopia.
3 Department of Anthropology and Archaeology, Authority for Research and Conservation of the Cultural Heritage, Ministry of Youth, Sports and Culture, Post Office Box 6686, Addis Ababa, Ethiopia.
4 Department of Physical Anthropology, Cleveland Museum of Natural History, 1 Wade Oval Drive, Cleveland, OH 44106, USA.
5 Department of Anthropology, School of Biomedical Sciences, Kent State University, Kent, OH 44240–0001, USA.
6 The University Museum, the University of Tokyo, Hongo, Bunkyo-ku, Tokyo 113-0033, Japan.
7 Earth Environmental Sciences Division, Los Alamos National Laboratory, Los Alamos, NM 87545, USA.
* To whom correspondence should be addressed. E-mail: timwhite@berkeley.edu
In 1871, Charles Darwin concluded that Africa was humanity’s most probable birth continent [(1), chapter 7]. Anticipating a skeptical reception of his placement of Homo sapiens as a terminal twig on the organic tree, Darwin lamented the mostly missing fossil record of early hominids (2). Following T. H. Huxley, who had hoped that "the fossilized bones of an Ape more anthropoid, or a Man more pithecoid" might be found by "some unborn paleontologist" [(3), p. 50], Darwin observed, "Nor should it be forgotten that those regions which are the most likely to afford remains connecting man with some extinct ape-like creature, have not as yet been searched by geologists." He warned that without fossil evidence, it was "useless to speculate on this subject" [(1), p. 199)].
Darwin and his contemporaries nonetheless sketched a scenario of how an apelike ancestor might have evolved into humans. That scenario easily accommodated fossil evidence then restricted to European Neandertals and Dryopithecus (a Miocene fossil ape). Javanese Homo erectus was found in the 1890s, followed by African Australopithecus in the 1920s. By the 1960s, successive grades of human evolution were widely recognized. Australopithecus comprised several Plio-Pleistocene small-brained species with advanced bipedality. This grade (adaptive plateau) is now widely recognized as foundational to more derived Homo.
Molecular studies subsequently and independently confirmed Huxley’s anatomically based phylogeny linking African apes and living humans (4). They also challenged age estimates of a human/chimpanzee divergence, once commonly viewed as exceeding 14 million years ago (Ma). The latter estimates were mostly based on erroneous interpretations of dentognathic remains of the Miocene fossil ape Ramapithecus, combined with the presumption that extant chimpanzees are adequate proxies for the last common ancestor we shared with them (the CLCA).
The phylogenetic separation of the lineages leading to chimpanzees and humans is now widely thought to have been far more recent. During the 1970s, discovery and definition of Australopithecus afarensis at Laetoli and Hadar extended knowledge of hominid biology deep into the Pliocene [to 3.7 Ma (5, 6)]. The slightly earlier (3.9 to 4.2 Ma) chronospecies Au. anamensis was subsequently recognized as another small-brained biped with notably large postcanine teeth and postcranial derivations shared with its apparent daughter species (7, 8). Late Miocene hominid fossils have been recently recovered from Ethiopia, Kenya, and Chad. These have been placed in three genera [Ardipithecus (9–12), Orrorin (13), and Sahelanthropus (14)]. They may represent only one genus (12, 15), and they challenge both savanna- and chimpanzee-based models (16) of hominid origins.
Continuing to build on fossil-free expectations traceable to Darwinian roots, some hold that our last common ancestors with African apes were anatomically and behaviorally chimpanzee-like (17), that extant chimpanzees can be used as "time machines" (18), and/or that unique features of Gorilla are merely allometric modifications to accommodate its great body mass. Thus, early Australopithecus has routinely been interpreted as "transitional" and/or a "locomotor missing link" (19, 20) between extant humans and chimpanzees. Bipedality is widely suggested to have arisen as an opportunistic, or even necessary, response to a drier climate and the expansion of savannas. These views have been challenged on paleontological and theoretical grounds (9, 21). However, without additional fossil evidence, the evolutionary paths of the various great apes and humans have remained shrouded.
In related papers in this issue (22–27), we describe in detail newly discovered and/or analyzed specimens of Ar. ramidus, including two individuals with numerous postcranial elements. All are dated to 4.4 Ma and come from the Middle Awash area of the Ethiopian Afar rift. Local geology and many associated fossils are also described (28–30). These new data jointly establish Ardipithecus as a basal hominid adaptive plateau preceding the emergence of Australopithecus and its successor, Homo. Inferences based on Ar. ramidus also facilitate understanding its precursors (22, 23, 27, 31). Here, we provide an integrated view of these studies and summarize their implications.
The Middle Awash. The Middle Awash study area contains a combined thickness of >1 km of Neogene strata. To date, these deposits have yielded eight fossil hominid taxa spanning the Late Miocene to Pleistocene (>6.0 to <0.08 Ma) (32, 33). Hominids make up only 284 of the 18,327 total cataloged vertebrate specimens. Spatially and chronologically centered in this succession, the Central Awash Complex (CAC) (28, 34) rises above the Afar floor as a domelike structure comprising >300 m of radioisotopically and paleomagnetically calibrated, sporadically fossiliferous strata dating between 5.55 and 3.85 Ma. Centered in its stratigraphic column are two prominent and widespread volcanic marker horizons that encapsulate the Lower Aramis Member of the Sagantole Formation (Fig. 1). These, the Gàala ("camel" in Afar language) Vitric Tuff Complex (GATC) and the superimposed Daam Aatu ("baboon" in Afar language) Basaltic Tuff (DABT), have indistinguishable laser fusion 39Ar/40Ar dates of 4.4 Ma. Sandwiched between the two tuffs are fossiliferous sediments averaging ~3 m in thickness and cropping out discontinuously over an arc-shaped, natural erosional transect of >9 km (28). The rich fossil and geologic data from these units provide a detailed characterization of the Pliocene African landscape inhabited by Ardipithecus.
|

We first surveyed the CAC during 1981 in attempts to understand the distribution of fossils within the region. We launched a systematic program of geological, geochronological, and paleontological investigation in 1992. Initial visits to the CAC’s northeastern flank documented abundant fossilized wood and seeds in the interval between the two tuffs. We collected and identified a highly fragmented sample of vertebrates, including abundant cercopithecid monkeys and tragelaphine bovids. The first hominid fossils were found at Aramis vertebrate paleontology locality 1 (ARA-VP-1) on 17 December 1992. Two initial seasons of stratigraphic and geochronological studies yielded 649 cataloged vertebrates, including a minimum number of 17 hominid individuals represented mostly by teeth (10).
Because of its content, the Lower Aramis Member became the focus of our paleontological efforts. Fourteen sublocalities within the original ARA-VP-1 locality were circumscribed and subjected to repeated collecting of all biological remains, based on multiple team crawls (35) across the eroding outcrops between 1995 and 2005. Analogous collections were made at adjacent localities (ARA-VP-6, -7, and -17), as well as at the eastern and western exposures of the Ardipithecus-bearing sedimentary units (KUS-VP-2 and SAG-VP-7) (KUS, Kuseralee Dora; SAG, Sagantole). The Lower Aramis Member vertebrate assemblage (table S1) now totals >6000 cataloged specimens, including 109 hominid specimens that represent a minimum of 36 individuals. An additional estimated 135,000 recovered fragments of bone and teeth from this stratigraphic interval are cataloged by locality and taxon as pooled "bulk" assemblages. Analogous samples were collected from the Lower Aramis Member on the eastern transect pole (SAG-VP-1, -3, and -6). Fossils from localities higher and lower in the local Middle Awash succession (7, 12, 32) and at nearby Gona (36) are reported elsewhere.
The ARA-VP-6/500 partial hominid skeleton. Bones of medium and large mammals were usually ravaged by large carnivores, then embedded in alluvial silty clay of the Lower Aramis Member. Once exposed by erosion, postdepositional destruction of the fossils by decalcification and fracture is typical. As a result, the larger vertebrate assemblage lacks the more complete cranial and postcranial elements typically recovered from other African hominid localities. The identification of larger mammals below the family level is therefore most often accomplished via teeth. The hominid subassemblage does not depart from this general preservational pattern (29).
There was consequently little initial hope that the stratigraphic interval between the two tuffs would yield crucially needed postcranial elements of Ardipithecus. The only relevant postcrania (arm elements) had come from slightly higher in the section in 1993 (10). However, on 5 November 1994, Y.H.S. collected two hominid metacarpal fragments (ARA-VP-6/500-001a and b) from the surface of an exposed silty clay ~3 m below the upper tuff (DABT), 54 m to the north of the point that had 10 months earlier yielded the Ardipithecus holotype dentition. Sieving produced additional hominid phalanges. The outcrop scrape exposed a hominid phalanx in situ, followed by a femur shaft and nearly complete tibia. Subsequent excavation during 1994 and the next field season (at a rate of ~20 vertical mm/day across ~3 m2) revealed >100 additional in situ hominid fragments, including sesamoids (Fig. 2 and table S2). Carnivore damage was absent.
|

The bony remains of this individual (ARA-VP-6/500) (Fig. 3) (37) are off-white in color and very poorly fossilized. Smaller elements (hand and foot bones and teeth) are mostly undistorted, but all larger limb bones are variably crushed. In the field, the fossils were so soft that they would crumble when touched. They were rescued as follows: Exposure by dental pick, bamboo, and porcupine quill probe was followed by in situ consolidation. We dampened the encasing sediment to prevent desiccation and further disintegration of the fossils during excavation. Each of the subspecimens required multiple coats of consolidant, followed by extraction in plaster and aluminum foil jackets, then additional consolidant before transport to Addis Ababa.
|
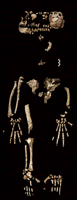
Pieces were assigned number suffixes based on recovery order. Back-dirt was weathered in place and resieved. The 1995 field season yielded facial fragments and a few other elements in northern and eastern extensions of the initial excavation. Further excavation in 1996 exposed no additional remains. Each fragment’s position, axial orientation, and dip were logged relative to a datum (strata here dip east at ~4° to 5°). A polygon representing the outer perimeter and vertical extent of the hominid fragment constellation (based on each bone’s center point) was demarcated by a carapace of limestone blocks cemented with concrete after excavation, then further protected by a superimposed pile of boulders, per local Afar custom.
The skeleton was scattered in typical Lower Aramis Member sediment (Fig. 2): fine-grained, massive, unslickensided, reddish-brown alluvial silty clay containing abundant decalcified root casts, fossil wood, and seeds. A 5- to 15-cm lens of poorly sorted sand and gravel lies immediately below the silty clay, and the spread of cranial parts to the north suggests that the bones of the carcass came to rest in a shallow swale on the floodplain.
There is no evidence of weathering or mammalian chewing on ARA-VP-6/500. Bony elements were completely disarticulated and lacked anatomical association. Many larger elements showed prefossilization fragmentation, orientation, and scatter suggestive of trampling. The skull was particularly affected, and the facial elements and teeth were widely scattered across the excavated area. Bioturbation tilted some phalanges and metacarpals at high dip angles (Fig. 2). A few postcrania of a large Aquila (eagle) and other birds were recovered during excavation, as were a few micromammals. No large-mammal remains (except isolated cercopithecid teeth and shaft splinters from a medium-to-large mammal limb bone) were associated. The cause of death is indeterminate. The specimen is judged to be female. The only pathology is a partially healed osteolytic lesion suggestive of local infection of the left proximal ray 5 pedal phalanx (ARA-VP-6/500-044).
Laboratory exposure and consolidation of the soft, crushed fossils were accomplished under binocular microscope. Acetone was applied with brushes and hypodermic needles to resoften and remove small patches of consolidant-hardened encasing matrix. Microsurgery at the interface between softened matrix and bone proceeded millimeter by submillimeter, rehardening each cleaned surface with consolidant after exposure. This process took several years. The freed specimens remain fragile and soft, but radiographic accessibility is excellent. Most restoration and correction for distortion were accomplished with plaster replicas or micro–computed tomography digital data to preserve the original fossils in their discovery state.
Environmental context. The Lower Aramis Member lacks any evidence of the hydraulic mixing that afflicts many other hominid-bearing assemblages. The unwarranted inference that early hominids occupied "mosaic habitats" (38) is often based on such mixed assemblages, so the resolution and fidelity of the Aramis environmental data sets are valuable. We estimate that the interval of time represented by the strata between the two tuffs at Aramis is <105 years, and perhaps just a few hundred or thousand years (28, 39). The lithology, thickness, taphonomic evidence, and similar age of the constraining marker horizons imply that geologically, the evidence can be viewed as "habitat time-averaged" (40). Indeed, we do not see notably different environmental indicators in the fossils or geologic or chemical data sampled vertically throughout the interval. The wealth of data allows a high-fidelity representation [sensu (41)] of the ecological community and environment inhabited by Ar. ramidus 4.4 Ma.
A variety of data indicate that the wooded biotope varied laterally across the Pliocene landscape (28–30). The hominid-bearing localities (centered on the ARA-VP-1 sublocalities) are rich in fossilized wood fragments, seeds, and animal fossils. Here, isotopic paleosol compositions indicate mostly wooded conditions (28). There was obviously more water at Aramis then (4.4 Ma)—supporting a much richer flora and fauna—than there is today. The higher water budget is possibly due to higher elevation during deposition (42) or to paleoclimatic factors such as a more continuous Pliocene El Niño effect (43). An abrupt transition occurs southeast of the SAG-VP-7 locality, where sedimentary, faunal, taphonomic, and isotopic data imply a more open rift-axial setting depauperate in faunal remains and lacking in primates, micromammals, and macrobotanical remains (29, 30).
Along the northern slope of the CAC, all localities of the Lower Aramis Member yielded tragelaphine bovids, monkeys, and other data indicative of more wooded conditions. Carbon isotopes from the teeth of five Ardipithecus individuals found here imply that they fed largely on C3 plants in woodlands and/or among the small patches of forests in the vicinity. We interpret the combined contextual data to indicate that Ar. ramidus preferred a woodland-to-forest habitat (29, 30) rather than open grasslands. This finding is inconsistent with hypotheses positing hominid origins via climate-driven savanna expansion.
Variation and classification. Initial (1994) description of the limited hominid sample from Aramis placed these remains in a newly discovered Australopithecus species interpreted as the most primitive then known (10). Subsequent recovery of the ARA-VP-6/500 skeleton showed that, relative to body size, its dentition was small, unlike Australopithecus. Strict cladistic practice required a new genus name for this sister taxon of Australopithecus, so the material was renamed as the new genus Ardipithecus in 1995, with the lack of megadonty added to the species diagnosis even as the partial skeleton’s excavation was still under way (44). Subsequent discovery of the earlier probable chronospecies Ar. kadabba in 1997 (11, 12) was followed by recovery of Orrorin in 2000 (13) and Sahelanthropus in 2001 (14). These Late Miocene fossils provide additional outgroup material useful in assessing the phylogenetic position of Ar. ramidus.
Only two adjacent Ethiopian study areas (the Middle Awash and Gona) have yielded confirmed remains of Ar. ramidus to date (7, 36). Neither has produced any evidence to reject a single species lineage as the source of the combined hominid sample from these Pliocene sites. We thus interpret the Lower Aramis Member hominid assemblage as a single taxon (22). Penecontemporary (~4.3 to 4.7 Ma) hominid remains from elsewhere are sparse (45, 46), and these are broadly compatible with the now expanded range of variation in Ar. ramidus (22, 23). Thus, although continental sampling is still obviously inadequate, describing hominid species diversity in this time frame (47) as "very bushy" seems unwarranted (48).
The amount of variation within the known Afar Ar. ramidus sample appears to be lower than typical for species of Australopithecus. This is probably due to a lesser degree of sexual dimorphism in Ardipithecus, combined with the narrow time window represented by the interval between the two Aramis tuffs. Skeletal dimorphism is notably difficult to assess, except in rare instances of geologically isochronous samples of a species lineage (e.g., A.L. 333 "first family") (49). For Ar. ramidus, the ARA-VP-6/500 skeleton (Figs. 3 and 4) provides a rare opportunity for guiding a probabilistic approach to sex attribution of conspecific fossils, relying on canines (22) and postcranially based estimates of body size (27). The implication is that there was broad overlap in body size between males and females of Ar. ramidus.
|
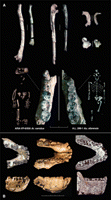
Cranial and dental anatomy. The Ar. ramidus skull (23) is very similar to the larger, more robust Sahelanthropus cranium (TM 266-01-60-1) from Chad, also interpreted as an early hominid (14, 50). Some of the differences are probably partly sex-related. Ar. ramidus shares with Sahelanthropus a small cranial capacity (300 to 350 cc) and considerable midfacial projection but a maxillo-premaxillary complex that is less prognathic than that of modern African apes [not necessarily a derived trait shared with Homo, in contrast with (51)]. The Ardipithecus and Sahelanthropus crania each lack a distinct post-toral sulcus, and both exhibit an anteriorly positioned posterior cranial base.
Most aspects of the craniofacial structure of Sahelanthropus/Ardipithecus are probably close to the African ape and hominid ancestral state. Gorilla and chimpanzee cranial morphologies, as well as their specialized dentitions, are clearly divergently derived (22). In Gorilla, enhanced facial size and prognathism occur in relation to larger general size and an increasing adaptation to herbivory and folivory. In Pan (also with enhanced prognathism), derived cranial form (including anterior basicranial lengthening) probably occurred as a part of enhanced terrestriality accompanied by elevated agonistic behavior and its anatomical correlates, such as tusklike canines (22, 23). The bonobo cranial base and Ardipithecus craniofacial structure may be less derived, but even the bonobo seems to be derived in its relatively small face and global dental reduction (22). This was probably at least in part due to decreased intraspecific aggression in the bonobo lineage after separation from the common chimpanzee lineage.
The superoinferiorly short but intermediately prognathic Ar. ramidus face lacks the broadening and anterior migration of the zygomaxillary area seen to varying degrees in species of Australopithecus. The primitive craniofacial pattern shared between Sahelanthropus and Ardipithecus suggests that the genus Australopithecus would later evolve a craniofacial structure capable of increased postcanine mastication consequent to an ecological breakout from wooded habitats, expanding its foraging into more open environments (7, 10).
The Ardipithecus dentition suggests omnivory (22). It exhibits none of the specializations seen among modern apes; neither the large incisors of Pongo or Pan nor the specialized molar morphology of Pongo, Pan, or Gorilla. Postcanine size relative to body size was slightly larger than in Pan but smaller than in Gorilla, Pongo, or (especially) Au. afarensis. Ar. ramidus molars overlap considerably with Pan in some measures of enamel thickness but differ in overall thickness and structure. Chimpanzee molars have a broad occlusal basin with locally thin enamel not seen in Ardipithecus. Pan molar morphology is probably an adaptation to crushing relatively soft and nonabrasive food items such as ripe fruits, while retaining some shearing capacities. The Ardipithecus dentition shows no strong signals of ripe-fruit frugivory, folivory-herbivory, or feeding on hard objects. Its macroscopic and microscopic wear patterns, as well as the low bunodont cusps with intermediate enamel thickness (22), suggest that its diet was not particularly abrasive but may have included some hard foods. It is consistent with a partially terrestrial, partially arboreal pattern of feeding in a predominantly wooded habitat.
Carbon isotopic evidence from the teeth of five Ar. ramidus individuals suggests that Ardipithecus and Australopithecus were distinct in dietary intake (30). "Robust" and "nonrobust" Australopithecus have enamel isotope values indicating a diet of more than 30% C4 plants, with variation ranging up to ~80% C4. In contrast, the known Ar. ramidus individuals vary only between ~10 and 25% C4, and thus also differ from Pan troglodytes, which prefers ripe fruit and is considered closer to a pure C3 feeder (30). Thus, Ardipithecus appears to have exploited a wider range of woodland resources than do chimpanzees, but without relying on the open biotope foods consumed by later Australopithecus.
Evolution of the canine/lower third premolar complex (C/P3) potentially illuminates social and reproductive behavior. The Ar. ramidus canine sample totals 21 Aramis individuals. Some are small fragments, but all show informative morphology and/or wear. All specimens are either morphologically similar to those from female apes or are further derived toward the later hominid condition (22). Morphological and metric variation in the sample is small. Functionally important sex-related size dimorphism is not apparent. There is no evidence of functional honing (planar facets on the mesiobuccal P3 or sharpened edges on the distolabial upper canine margin). The largest, presumably male, specimens are as morphologically derived as the smallest, showing that dimorphic canine morphology was virtually absent in these hominids by 4.4 Ma. Furthermore, a juvenile probable male lacks the delayed canine eruption seen in chimpanzees, approximating the Au. anamensis and Au. afarensis conditions and indicating that the canine was not an important component of adult sociobehavioral relationships.
The differential status of upper versus lower canine morphology is informative. In Ar. ramidus, the lower canines retain modally more apelike morphology than do the uppers, and, in contradistinction to other anthropoids, the height of the maxillary canine crown is lower than that of the mandibular (22). This relationship is opposite that seen in great apes and cercopithecids, whose upper canine dominance is exaggerated, particularly in males of dimorphic species. In these primates, upper canine projection and prominence function in both weaponry and display. The Ar. ramidus canines are metrically and morphologically derived in the direction of later hominids, and we hypothesize that reduction and alteration of upper canine size and shape in this and earlier hominid species are related to changes in social behaviors (22, 31).
The canines of Sahelanthropus, Orrorin, and Ar. kadabba are broadly equivalent to those of Ar. ramidus in size and function. However, the upper canines of Late Miocene hominids exhibit a subtle but distinctly more primitive morphology than their Ar. ramidus homologs, potentially including occasional residual (female ape–like) honing as part of their variation (12, 15). This suggests that upper canine prominence was reduced through the Late Miocene and Early Pliocene. In contrast, the C/P3 complex of the last common ancestor of hominids and chimpanzees probably had a moderate level of canine dimorphism combined with functional honing. This was subsequently generally retained in P. paniscus and enhanced in P. troglodytes.
Body size and dimorphism. The partial skeleton ARA-VP-6/500 is identified as female based on probability assessments of canine size (its canines are among the smallest of those of 21 available individuals) (22). This interpretation is corroborated by its small endo- and exocranial size, as well as its superoinferiorly thin supraorbital torus (23). Bipedal standing body height for the ARA-VP-6/500 individual is estimated at approximately 120 cm, and body mass at ~50 kg (27). Although actual body mass may vary considerably in relation to skeletal size, this is a large female body mass.
Of the Ar. ramidus postcranial elements, the humerus represents the largest minimum number of individuals (seven). ARA-VP-6/500 does not preserve a humerus, but detailed comparisons suggest that its forelimb was ~2 to 8% larger in linear dimensions than the partial forelimb skeleton ARA-VP-7/2 (24, 27), which does include a humerus. This would make ARA-VP-6/500 either the second- or third-largest of eight individuals within the Aramis humeral sample. The combined evidence suggests that Ardipithecus skeletal body size was nearly monomorphic, and less dimorphic than Australopithecus, as estimated from template bootstrapping (49). Most likely, Ardipithecus exhibited minimal skeletal body size dimorphism, similar to Pan, consistent with a male-bonded social system, most likely a primitive retention from the CLCA condition (31). With its subsequent commitment to terrestrial bipedality, Australopithecus probably enhanced female cooperation and group cohesion, thus potentially reducing female body size, whereas male size increased in response to predation pressure, probably elevated by expanding niche breadth.
Postcranial biology and locomotion. Regardless of whether the Afar Ar. ramidus population represents a hominid relict or a lineal ancestor, this taxon’s biology resolves fundamental evolutionary questions persisting since Darwin. Its substantially primitive postcranial anatomy appears to signal a grade-based difference from later Australopithecus. The challenge of understanding its evolutionary and functional implications required a nontraditional approach. Without testable hypotheses of underlying gene-based developmental mechanisms, many paleoanthropological analyses have been adaptationist (52) and/or purely numerically discriminatory. Therefore, wherever possible, in the accompanying postcranial papers (24–27) we restrict hypotheses to those that can be formulated consistent with putative selection acting on cascades of modular-based positional information, especially when these can be potentially grounded in known anabolic mechanisms. This approach is summarized elsewhere (53, 54) and in supporting online material text S1.
The upper pelvis of Ar. ramidus presents a contrast to its primitive hand, foot, and limbs. The ilia are abbreviated superoinferiorly and sagittally oriented but broad mediolaterally, so much so that the anterior inferior iliac spine has become a separate growth site, as in all later hominids. The pubic symphyseal face is quite short. A slight sciatic notch is present, although ischial structure was similar to that of extant African apes. This suggests that pattern-formation shifts for bipedality were only partly realized in Ar. ramidus. These changes may have culminated a long period of facultative bipedality hinted at by isolated postcranial elements from the probable chronospecies Ar. kadabba (12) and other Late Miocene forms (13, 14).
Paramount among the retained primitive characters of the Ar. ramidus hindlimb is a fully abductable first ray (hallux, or great toe), but in combination with elements of a robust plantar substructure that stabilized the foot during heel- and toe-off. Although it was still a highly effective grasping organ, the foot of Ar. ramidus also maintained propulsive capacity long since abandoned by extant great apes (in which greater opposition between the hallux and lateral rays evolved, i.e., a more handlike conformation than in Ar. ramidus) (26).
Other defining and notably primitive characters include a moderately elongate mid-tarsus, a robust lateral peroneal complex in which muscles of the lateral compartment performed substantial plantarflexion, and a primitive (flexion-resistant) geometric configuration of the lateral metatarsal bases. Thus, the Ar. ramidus foot is an amalgam of retained primitive characters as well as traits specialized for habitual bipedality, such as the expanded second metatarsal base that anchored plantarflexion during heel- and toe-off. Many of the foot’s primary adaptations to fulcrumation are probable retentions from the gorilla/chimpanzee/human last common ancestor (GLCA), but these have been eliminated in apes, presumably for vertical climbing.
The ARA-VP-6/500 radius/tibia ratio is 0.95, as in generalized above-branch quadrupeds such as macaques and Proconsul (an Early Miocene ape) (27). Its intermembral index (the ratio of forelimb length to hindlimb length) is also similar to those of above-branch quadrupeds. These facts suggest that African apes experienced both forelimb elongation and hindlimb reduction, whereas hominid proportions remained largely unchanged until the dramatic forearm shortening and hindlimb elongation of Plio-Pleistocene Homo.
These primitive proportions are consistent with virtually all other aspects of the Ar. ramidus skeleton. The inferred locomotor pattern combined both terrestrial bipedality and arboreal clambering in which much weight was supported on the palms. The hand phalanges are elongate relative to those of Proconsul, but metacarpals (Mc) 2 to 5 remained primitively short and lacked any corporal modeling or adaptations typical of knuckle-walking (24). Moreover, the virtually complete wrist of ARA-VP-6/500 (lacking only the pisiform) exhibits striking adaptations for midcarpal dorsiflexion (backward deflection of the dorsum of the hand), consistent with a highly advanced form of arboreal palmigrady. In addition, substantial metacarpal-phalangeal dorsiflexion is indicated both by moderate dorsal notching of the Mc2 to -5 heads and by marked palmar displacement of the capitate head. Together these must have permitted dorsiflexion of the wrist and hand to a degree unparalleled in great apes.
The Ar. ramidus elbow joint provided full extension but lacks any characters diagnostic of habitual suspension. Ulnar withdrawal was complete and the thumb moderately robust, with indications of a distinct and fully functional flexor pollicis longus tendon. The hamate’s hamulus permitted substantial metacarpal motion for opposition against the first ray. The central joint complex (Mc2/Mc3/capitate/trapezoid) exhibits none of the complex angular relationships and marked syndesmotic reinforcement seen in extant apes. Together, these retained primitive characters, unlike their homologs in highly derived African apes, imply that the dominant locomotor pattern of the GLCA was arboreal palmigrady rather than vertical climbing and/or suspension (orthogrady). Another strong inference is that hominids have never knuckle-walked (26).
The extraordinary forelimb of Ar. ramidus, in combination with its limb proportions and likely primitive early hominid lumbar column (55), casts new light on the evolution of the lower spine. The traditional interpretation has been that the lumbar transverse processes became dorsally relocated as the lumbar column reduced in length. The data from Ar. ramidus imply that ulnar withdrawal was not a suspensory adaptation but was instead an enhancement of distal forelimb maneuverability that accompanied profound changes in the shoulder. Spinal column invagination appears to have been an integral part of thoracic restructuring to increase shoulder joint laterality, thereby enhancing forelimb mobility for advanced arboreal quadrupedalism, especially careful climbing and bridging. A still primitive deltoid complex in both Ar. ramidus and Asian ancestral apes (e.g., Sivapithecus) now becomes more understandable. A predominantly Sharpey’s fiber deltoid insertion can be viewed as a retention in above-branch quadrupeds that only later became modified for suspension (separately) in extant African and Asian apes.
The adoption of bipedality and its temporal association with progressive canine reduction and loss of functional honing now constitute the principal defining characters of Hominidae. The orthograde positional behaviors of hominids and apes were thus acquired in parallel, generated by early bipedal progression in the former and suspension and vertical climbing in the latter. Overall, Ar. ramidus demonstrates that the last common ancestors of humans and African apes were morphologically far more primitive than anticipated, exhibiting numerous characters reminiscent of Middle and Early Miocene hominoids. This reinforces what Huxley appreciated in 1860: "the stock whence two or more species have sprung, need in no respect be intermediate between those species" [(56), p. 568].
Ardipithecus and the great apes. Ar. ramidus illuminates several collateral aspects of hominoid evolution. Despite the demise of Ramapithecus as a putative hominid ancestor, at least one Eurasian Miocene ape, Ouranopithecus, has been suggested as being phyletically related to later African hominids (57), whereas another, Dryopithecus, is often considered an alternative sister taxon of the hominid and African ape clade (58). Ardipithecus effectively falsifies both hypotheses. Ar. ramidus lacks the derived characters of Ouranopithecus associated with postcanine enlargement and relative canine reduction while still providing a primitive morphological substrate for the emergence of Australopithecus. The new perspective that Ar. ramidus offers on hominoid postcranial evolution strongly suggests that Dryopithecus acquired forelimb adaptations to suspensory behaviors independently from African apes. Ar. ramidus suggests that these Eurasian forms were too derived to have been specially related to either the hominid or extant African ape clades. Moreover, the remarkably primitive postcranium of potential Pongo ancestors (e.g., Sivapithecus), coupled with what is now evidently widespread homoplasy in extant hominoids, suggests that the Pongo clade was established even before the first dispersal events of large-bodied apes from Africa into Eurasia, shortly after docking of the Afro-Arabian and Eurasian plates at ~18 Ma (59).
An additional implication of Ar. ramidus stems from its demonstration that remarkable functional and structural similarities in the postcrania of Pongo and the African apes have evolved in parallel, as have those of Pan and Gorilla (27). Until now, a myriad of characters shared among the extant African apes were presumed to have been present also in ancestral hominids (because they were presumed to have been the ancestral state) (60). However, it now appears that many of these putative shared primitive characteristics have evolved independently. This highlights the alacrity with which similar anatomical structures can emerge, most likely by analogous selection operating on homologous genomes. The same genetic pathways can be repeatedly and independently coopted, resulting in convergent adaptations (61). Recent work on gene expression demonstrates that there are also multiple pathways that can produce similar but independently derived anatomical structures (62).
Work on deep homology shows that parallel evolution "must be considered a fact of life in the phylogenetic history of animals" [(63), p. 822]. This is also seen in more terminal branches; for example, during the past two million years of stickleback fish evolution (64). Such evolvability and parallelism are even suggested for the catarrhine dentition (65). Ar. ramidus reveals an excellent example of this phenomenon within the African ape-hominid clade by demonstrating the striking reoccurrence of syndesmotic fixation of the central joint complexes in hominoid wrists adapted to suspensory locomotion (including not only those of Pan and Gorilla but also those of Pongo and, partially, Dryopithecus). Such observations on very different evolutionary scales all caution against indiscriminant reliance on raw character states to assess phylogeny. A consideration of wider patterns of manifestations of such adaptive evolution, not only in character constellations but also in their evolutionary context, may be needed to tease apart homology and homoplasy. A far more complete fossil record will be needed to accomplish such a goal.
Such considerations also bear on current estimates of the antiquity of the divergence between the human and chimpanzee clades. Many such estimates, suggesting striking recency, have become widely accepted because of the presumed homology of human and African ape morphologies (60). This obtains despite the recognition that broad assumptions about both the regularity of molecular change and the reliability of calibration dates required to establish such rates have strong limitations (66, 67). The homoplasy now demonstrated for hominoids by Ar. ramidus provides fair warning with respect to such chronologies, especially those currently used to calibrate other divergence events, including the split times of New and Old World monkeys, hylobatids, and the orangutan. The sparseness of the primate fossil record affecting these estimates is now compounded by the dangers posed by convergences perceived as homologies. Such difficulties are further exacerbated by newly recognized complexities in estimating quantitative degrees of genetic separation (66–68). In effect, there is now no a priori reason to presume that human-chimpanzee split times are especially recent, and the fossil evidence is now fully compatible with older chimpanzee-human divergence dates [7 to 10 Ma (12, 69)] than those currently in vogue (70).
Hominid phylogenetics. The expanded Ar. ramidus sample allows more detailed consideration of early hominid phylogenetics. The placement of Ardipithecus relative to later hominids can be approached by using modern and Miocene apes as the outgroup. An earlier cladistic study of this kind concluded that Ar. ramidus was the sister taxon of all later hominids (71). A more recent assessment of Ar. ramidus dental characters came to the same conclusion (7). In these analyses, a suite of derived features and character complexes exclusively aligning Ar. ramidus with Australopithecus was identified, but these were based on comparatively limited anatomical elements. The Ar. ramidus characters reported here, combined with those from Gona (36), allow a more complete analysis that clarifies the relationships among early hominid taxa.
Parsimony-based cladistic analyses are useful in deciphering relationships within the hominid family tree, despite their shortcomings (72, 73). The distribution of characters identified in Table 1 clearly shows that Ar. ramidus is derived relative to all known Late Miocene fossils attributed to the hominid clade. The earlier and more primitive probable chronospecies Ar. kadabba is found in 5.5- to 5.7-million-year-old deposits a mere 22 km west of Aramis, consistent with local (and perhaps regional) phyletic evolution. Its limited known elements are similar to those of other Late Miocene hominids in Kenya and Chad (12–14).
|
Comparatively few features of Ar. ramidus are derived relative to these earlier hominids, although many body parts of the latter are still unrepresented. There are no apparent features sufficiently unique to warrant the exclusion of Ar. ramidus as being ancestral to Australopithecus (74), and a greatly expanded set of shared derived characters now links Ar. ramidus with later members of the hominid clade. Table 1 identifies some of the most important. This pattern robustly falsifies earlier assessments that the Aramis fossils represent an ancestral chimpanzee (13, 75). These results are suggestive of a cohesive hominid evolutionary grade preceding Australopithecus (currently >6.0 to 4.2 Ma). By priority, the name Ardipithecus may encompass other named genera at this adaptive plateau (12, 15).
The question of whether Ar. ramidus is ancestral to later hominids is moot for some cladists because they consider ancestors inherently unrecognizable and therefore recognize only sister taxa (76). The fossils reported here make it even more obvious that Ar. ramidus is the cladistic sister to Australopithecus/Homo because it shares primitive characters with earlier hominids and apes but at the same time exhibits many important derived characters that are shared exclusively only with later hominids (Table 1).
Species-level phylogenetics are more difficult to discern given the sparse geographic and temporal distribution of available fossils (Fig. 5). Primitive characters seen in Ar. ramidus persist most markedly in its apparent (but still poorly sampled) sister species Au. anamensis and, to a lesser degree, in Au. afarensis. The known dental and mandibular remains of Au. anamensis are temporally and morphologically intermediate between those of Ar. ramidus and Au. afarensis, with variation that overlaps in both directions. Its constellation of primitive and derived features of the mandible, CP3 complex, lower dm1 (lower first deciduous molar), and postcanine dentition lends support to the hypothesis of an evolutionary sequence of Ar. ramidus →Au. anamensis →Au. afarensis (7, 8, 77). Circumstantial evidence supporting this hypothesis is the temporal and geographic position of Ar. ramidus directly below the first known appearance of Au. anamensis within the Middle Awash succession. Here, these taxa are stratigraphically superimposed in the same succession, separated by ~80 vertical meters representing ~200,000 to 300,000 years (7). Au. afarensis appears later in the same sequence [3.4 Ma, at Maka (53)].
|
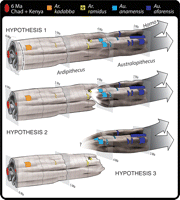
Therefore, at one end of a spectrum of phylogenetic possibilities, Ar. ramidus may have been directly ancestral to the more derived chronospecies pair Au. anamensis →Au. afarensis across the full (still unknown, presumably African) species range (7, 8, 77) (Fig. 5A). Although Au. afarensis is well represented in craniodental remains and postcrania, its apparent earlier chronospecies Au. anamensis is still woefully underrepresented in both, and because Ar. ramidus is so far known only from limited time horizons and locations, its last appearance, date, and potential relationship to these later taxa are still indeterminate. Given the dramatic differences in postcranial anatomy seen in later Australopithecus and hinted at in known Au. anamensis, it seems likely that a major adaptive shift marked the Ardipithecus-to-Australopithecus transition (whenever and wherever the transition might have occurred and whatever its population dynamics). This transition may not have occurred through pan-specific phyletic evolution (Fig. 5A). Figure 5 presents two other phylogenetic hypotheses that are also, at present, impossible to falsify.
If diagnostic contemporary fossils of Au. anamensis are someday found in rocks of >4.4 Ma, the hypothesis that the Afar population of Ar. ramidus is the phyletic ancestor of Au. anamensis (Fig. 5A, B) would be falsified. In such an eventuality, Aramis Ar. ramidus would represent a persisting relict population of the mother species (Fig. 5C). Given the lack of relevant fossils, it is currently impossible to determine whether there was a geologically rapid phyletic transition between Ardipithecus and Australopithecus in the Middle Awash or elsewhere. Nevertheless, the morphological and ecological transition between these two adaptive plateaus is now discernible.
Ardipithecus and Australopithecus. For Darwin and Huxley, the basic order in which human anatomies, physiologies, and behaviors were assembled through time was unknown—indeed unknowable—without an adequate fossil record. They were forced to employ extant ape proxies instead. The latter are now shown to be derived in ways unrelated to the evolution of hominids.
The Aramis fossils help clarify the origin of the hominid clade (27, 31), and reveal some paleobiological dimensions of the first hominid adaptive plateau (Ardipithecus). The primitive characters of Ar. ramidus simultaneously provide a new perspective on the evolutionary novelties of Australopithecus.
Even in the wake of the Aramis and Gona discoveries, the morphological envelopes, phylogenetic relationships, and evolutionary dynamics of early hominid species remain incompletely understood (Fig. 5). However, the paleobiology of Ar. ramidus—even when viewed through its geographically and temporally restricted Afar samples—now reveals that the basal hominid adaptive plateau comprised facultatively bipedal primates with small brains, reduced nonhoning canines, unspecialized postcanine dentitions, and arboreally competent limb skeletons. Their ecological niche(s) were probably more restricted—and their geographic distribution(s) possibly smaller and more disjunct—than those of later hominid species and genera.
The derived postcranial elements of Australopithecus provide a strong contrast to their more primitive homologs in Ardipithecus (78). Relative to the generalized anatomy of the latter, the highly evolved specializations of the foot, ankle, knee, pelvis, wrist, and hand of Au. afarensis (79–81) indicate that this species lineage had largely abandoned locomotion in the arboreal canopy (and its resources).
Given the strong selection predicted to have been associated with the emergence of new ranging and feeding patterns in Australopithecus, the transition from Ardipithecus to Australopithecus could have been rapid, and anatomically particularly so in hindlimb structure. The forelimb (especially the hand) was probably under less intensive selection. It is possible that modification of general cis-regulatory pathways may have generated the striking and novel morphology of the hindlimb, especially the foot, because the autopod seems to be the most morphologically compliant to such mechanisms of modification. The dentognathic shifts could have been more gradational, whatever the mode of phylogenesis.
Homo and Australopithecus are the only primates with nongrasping feet, and this particular transformation was probably far-reaching, with consequences for key behavioral constancies in higher primates related to arboreal feeding and nesting. Without stabilizing selection for Ardipithecus-like arboreal capacities involving slow and careful climbing, the foot, pelvis, and thigh would have experienced directional selection to optimize bipedal locomotion during prolonged walking (also in more limited running bouts). With expanded ranging and social adaptations associated with terrestrial feeding in increasingly open environments, the transition could have been profound, but probably rapid, and therefore difficult to probe paleontologically.
One possible dynamic of an Ardipithecus-to-Australopithecus transition would have involved microevolution within a deme or regional group of demes. Being more ecologically flexible, the derived, potentially speciated populations would have undergone rapid range expansion, perhaps even encountering relict Ardipithecus populations. Unfortunately, the phylogeographic details remain obscure given the poor spatial and temporal resolution of the current fossil record (Fig. 5). This provides a strong incentive for pursuing that record by actively increasing sampling of sediments from different African basins with dates between ~5 and ~3.5 Ma.
Currently, Australopithecus appears relatively abruptly in the fossil record at about 4.2 Ma. Relative to Ar. ramidus, available early Australopithecus is now revealed to have been highly derived: a committed biped with slightly enlarged brain, a nongrasping arched foot, further derived canines, substantially specialized postcanine teeth with thick molar enamel, and expanded ecological tolerances and geographic ranges. It is widely recognized that this is the adaptive plateau antecedent to Homo, which is now definable as the third such major adaptive shift in human evolution. Commitment to the terrestrial ranging behaviors of Australopithecus well before the Pleistocene appear to have catalyzed the emergence of what must have been even more highly specialized social and ecological behaviors remarkably elaborated in descendant Homo—the ultimate global primate generalist.
Conclusions. Besides hominids, the only apes to escape post-Miocene extinction persist today as relict species, their modern distributions centered in forested refugia. The markedly primitive Ar. ramidus indicates that no modern ape is a realistic proxy for characterizing early hominid evolution—whether social or locomotor—as appreciated by Huxley. Rather, Ar. ramidus reveals that the last common ancestor that we share with chimpanzees (CLCA) was probably a palmigrade quadrupedal arboreal climber/clamberer that lacked specializations for suspension, vertical climbing, or knuckle-walking (24–27). It probably retained a generalized incisal/postcanine dentition associated with an omnivorous/frugivorous diet less specialized than that of extant great apes (22, 23). The CLCA probably also combined moderate canine dimorphism with minimal skull and body size dimorphism (22, 23), most likely associated with relatively weak male-male agonism in a male philopatric social system (22, 23, 31).
Ardipithecus reveals the first hominid adaptive plateau after the CLCA. It combined facultative terrestrial bipedality (25, 26) in a woodland habitat (28–30) with retained arboreal capabilities inherited from the CLCA (24–27). This knowledge of Ar. ramidus provides us, for the first time, with the paleobiological substrate for the emergence of the subsequent Australopithecus and Homo adaptive phases of human evolution. Perhaps the most critical single implication of Ar. ramidus is its reaffirmation of Darwin’s appreciation: Humans did not evolve from chimpanzees but rather through a series of progenitors starting from a distant common ancestor that once occupied the ancient forests of the African Miocene.
www.sciencemag.org/cgi/content/full/326/5949/64/DC1
SOM Text
Tables S1 and S2
References
- 1. C. Darwin, The Descent of Man, and Selection in Relation to Sex (John Murray, London, 1871).
- 2. We here consider Hominidae to include modern humans and all taxa phylogenetically closer to humans than to Pan (common chimpanzee and bonobo), that is, all taxa that postdate the split between the lineage leading to modern humans and the lineage that led to extant chimpanzees.
- 3. T. H. Huxley, Evidence as to Man’s Place in Nature (London, 1863).
- 4. V. M. Sarich, A. C. Wilson, Proc. Natl. Acad. Sci. U.S.A. 58, 142 (1967).
[Free Full Text] - 5. D. C. Johanson, M. Taieb, Y. Coppens, Am. J. Phys. Anthropol. 57, 373 (1982). [CrossRef] [Web of Science] [Medline]
- 6. M. D. Leakey et al., Nature 262, 460 (1976). [CrossRef]
- 7. T. D. White et al., Nature 440, 883 (2006). [CrossRef] [Medline]
- 8. W. H. Kimbel et al., J. Hum. Evol. 51, 134 (2006). [CrossRef] [Web of Science] [Medline]
- 9. G. WoldeGabriel et al., Nature 371, 330 (1994). [CrossRef] [Medline]
- 10. T. D. White, G. Suwa, B. Asfaw, Nature 371, 306 (1994). [CrossRef]
- 11. Y. Haile-Selassie, Nature 412, 178 (2001). [CrossRef]
- 12. Y. Haile-Selassie, G. Suwa, T. D. White, in Ardipithecus kadabba: Late Miocene Evidence from the Middle Awash, Ethiopia, Y. Haile-Selassie, G. WoldeGabriel, Eds. (University of California, Berkeley, CA, 2009), pp. 159–236.
- 13. B. Senut et al., Comptes Rendus de l’Academie des Sciences, Series IIA: Earth and Planetary Science 332, 137 (2001). [CrossRef]
- 14. M. Brunet et al., Nature 418, 145 (2002). [CrossRef]
- 15. Y. Haile-Selassie, G. Suwa, T. D. White, Science 303, 1503 (2004).
[Abstract/Free Full Text] - 16. J. Moore, in Great Ape Societies, W. C. McGrew et al., Eds. (Cambridge Univ. Press, Cambridge, 1996), pp. 275–292.
- 17. B. G. Richmond, D. R. Begun, D. S. Strait, Yearb. Phys. Anthropol. 44, 70 (2001). [Web of Science]
- 18. R. Wrangham, D. Pilbeam, in All Apes Great and Small, Volume 1: African Apes, B. Galdikas et al., Eds. (Kluwer Academic/Plenum, New York, 2001), pp. 5–17.
- 19. J. T. Stern, R. L. Susman, Am. J. Phys. Anthropol. 60, 279 (1983). [CrossRef] [Web of Science] [Medline]
- 20. J. T. Stern, Evol. Anthropol. 9, 113 (2000). [CrossRef] [Web of Science]
- 21. B. Latimer, in Origines de la Bipedie chez les Hominides, B. Senut, Y. Coppens, Eds. (Editions CNRS, Paris, 1991), pp. 169–176.
- 22. G. Suwa et al., Science 326, 69 (2009).
[Abstract/Free Full Text] - 23. G. Suwa et al., Science 326, 68 (2009).
[Abstract/Free Full Text] - 24. C. O. Lovejoy et al., Science 326, 70 (2009).
[Abstract/Free Full Text] - 25. C. O. Lovejoy et al., Science 326, 71 (2009).
[Abstract/Free Full Text] - 26. C. O. Lovejoy et al., Science 326, 72 (2009).
[Abstract/Free Full Text] - 27. C. O. Lovejoy et al., Science 326, 73 (2009).
[Abstract/Free Full Text] - 28. G. WoldeGabriel et al., Science 326, 65 (2009).
[Abstract/Free Full Text] - 29. A. Louchart et al., Science 326, 66 (2009).
[Abstract/Free Full Text] - 30. T. D. White et al., Science 326, 67 (2009).
[Abstract/Free Full Text] - 31. C. O. Lovejoy, Science 326, 74 (2009).
[Abstract/Free Full Text] - 32. H. Gilbert, B. Asfaw (Eds.), Homo erectus: Pleistocene Evidence from the Middle Awash, Ethiopia (Univ. California Press, Berkeley, CA, 2008).
- 33. Y. Haile-Selassie, G. WoldeGabriel (Eds.), Ardipithecus kadabba: Late Miocene Evidence from the Middle Awash, Ethiopia (Univ. California Press, Berkeley, California, 2009).
- 34. P. R. Renne, G. WoldeGabriel, W. K. Hart, G. Heiken, T. D. White, Geol.
Soc. Am. Bull. 111, 869 (1999).
[Abstract/Free Full Text] - 35. In 1994, the Middle Awash project instituted "crawls" of sedimentary outcrop between the GATC and DABT to collect all available fossil material. Crawls were generally upslope in direction, done by teams of 5 to 15 collectors who crawled the surface on hands and knees, shoulder to shoulder, collecting all fossilized biological materials between a prescribed pair of taut nylon cords. Surfaces were repeatedly collected with this technique, invariably resulting in successively depressed specimen recovery numbers in subsequent field seasons.
- 36. S. Semaw et al., Nature 433, 301 (2005). [CrossRef] [Web of Science] [Medline]
- 37. No surface or in situ fragments of the ARA-VP-6/500 specimen are duplicate anatomical elements. Only 7.3% of 136 total pieces (table S2) were surface recoveries at the excavation site. All other pieces were excavated in situ. Preservation is identical across the entire recovered set. There is no evidence of multiple maturational ages among the 136 pieces, and many of them conjoin. Given the close stratigraphic and spatial association (Fig. 2), and given no evidence of any other individual from the carefully excavated spatiostratigraphic envelope, we conclude that the parts of the ARA-VP-6/500 specimen represent a single individual’s disarticulated skeleton.
- 38. S. Elton, J. Anat. 212, 377 (2008). [CrossRef] [Web of Science] [Medline]
- 39. A. K. Behrensmeyer, Paleobiology 8, 211 (1981). [Web of Science]
- 40. S. M. Kidwell, K. W. Flessa, Annu. Rev. Earth Planet. Sci. 24, 433 (1996). [CrossRef] [Web of Science]
- 41. D. Western, A. K. Behrensmeyer, Science 324, 1061 (2009).
[Abstract/Free Full Text] - 42. R. Pik, B. Marty, J. Carignan, J. Lavé, Earth Planet. Sci. Lett. 215, 73 (2003). [CrossRef] [Web of Science]
- 43. A. V. Fedorov et al., Science 312, 1485 (2006).
[Abstract/Free Full Text] - 44. T. D. White, G. Suwa, B. Asfaw, Nature 375, 88 (1995). [CrossRef] [Medline]
- 45. A. Hill, S. Ward, Yearb. Phys. Anthropol. 31, 49 (1987). [CrossRef]
- 46. M. G. Leakey, J. M. Harris, Eds., Lothagam: The Dawn of Humanity in Eastern Africa (Columbia Univ. Press, New York).
- 47. J. Kappelman et al., Nature 376, 558 (1995). [Web of Science] [Medline]
- 48. T. D. White, in The Paleobiological Revolution: Essays on the Growth of Modern Paleontology, D. Sepkoski, M. Ruse, Eds. (Univ. of Chicago Press, Chicago, 2009), pp. 121–148.
- 49. P. L. Reno, R. S. Meindl, M. A. McCollum, C. O. Lovejoy, Proc. Natl.
Acad. Sci. U.S.A. 100, 9404 (2003).
[Abstract/Free Full Text] - 50. M. Brunet et al., Nature 434, 752 (2005). [CrossRef] [Medline]
- 51. B. Wood, Nature 418, 133 (2002). [CrossRef] [Medline]
- 52. S. J. Gould, R. C. Lewontin, Proc. R. Soc. London Ser. B. 205, 147 (1979).
- 53. C. O. Lovejoy, R. S. Meindl, J. C. Ohman, K. G. Heiple, T. D. White, Am. J. Phys. Anthropol. 119, 97 (2002). [CrossRef] [Web of Science] [Medline]
- 54. C. O. Lovejoy, M. J. Cohn, T. D. White, Proc. Natl. Acad. Sci. U.S.A.
96, 13247 (1999).
[Abstract/Free Full Text] - 55. M. A. McCollum et al., J. Exp. Zool. B Mol. Dev. Evol. 312, published online 17 August 2009; 10.1002/jez.b.21316.
- 56. T. H. Huxley, Westminster Rev. 73, 541 (1860).
- 57. L. de Bonis, G. D. Koufos, Evol. Anthropol. 3, 75 (1994). [CrossRef]
- 58. D. R. Begun, Anthropol. Sci. 113, 53 (2005). [CrossRef]
- 59. J. Palfy et al., Earth Planet. Sci. Lett. 258, 160 (2007). [CrossRef]
- 60. D. Pilbeam, N. Young, C. R. Palevol 3, 305 (2004). [CrossRef] [Web of Science]
- 61. D. S. Stern, V. Orgogozo, Science 323, 746 (2009).
[Abstract/Free Full Text] - 62. M. D. Shapiro, M. A. Bell, D. M. Kingsley, Proc. Natl. Acad. Sci. U.S.A.
103, 13753 (2006).
[Abstract/Free Full Text] - 63. N. Shubin, C. Tabin, S. Carroll, Nature 457, 818 (2009). [CrossRef] [Web of Science] [Medline]
- 64. P. F. Colosimo et al., Science 307, 1928 (2005).
[Abstract/Free Full Text] - 65. L. Hlusko, G. Suwa, R. T. Kono, M. C. Mahaney, Am. J. Phys. Anthropol. 124, 223 (2004). [CrossRef] [Web of Science] [Medline]
- 66. M. J. F. Pulquerio, R. A. Nichols, Trends Ecol. Evol. 22, 180 (2007). [CrossRef] [Medline]
- 67. N. Elango, J. W. Thomas, S. V. Yi, Proc. Natl. Acad. Sci. U.S.A. 103,
1370 (2006).
[Abstract/Free Full Text] - 68. R. J. Britten, Proc. Natl. Acad. Sci. U.S.A. 99, 13633 (2002).
[Abstract/Free Full Text] - 69. G. Suwa, R. T. Kono, S. Katoh, B. Asfaw, Y. Beyene, Nature 448, 921 (2007). [CrossRef] [Medline]
- 70. N. Patterson, D. J. Richter, S. Gnerre, E. S. Lander, D. Reich, Nature 441, 1103 (2006). [CrossRef] [Medline]
- 71. D. S. Strait, F. E. Grine, J. Hum. Evol. 47, 399 (2004). [CrossRef] [Web of Science] [Medline]
- 72. E. Trinkaus, Am. J. Phys. Anthropol. 83, 1 (1990). [CrossRef] [Web of Science] [Medline]
- 73. For example, it has been noted that these methods fail to accurately resolve relationships of modern hominoid species without sufficient intermediate forms from a fossil record (71).
- 74. Enamel thickness of Ar. ramidus molars ranges largely from what would traditionally be termed "intermediate thin" to "intermediate thick" categories. Lacking the derived thickness pattern of Pan, it forms a suitable ancestral condition for later Australopithecus. The ubiquitous single-rooted lower fourth premolar (P4) in known Aramis and Gona Ar. ramidus is notable, but this is also a known variation of Au. anamensis and A. afarensis. Judging from the clear dominance of double-rooted lower P4’s in Au. afarensis (and thereafter an increasing robusticity of the roots themselves in Australopithecus), either there was selection for larger, more complex premolar root systems or such morphologies emerged as pleiotropy of postcanine enhancement. Without such selection, Ar. ramidus as a species probably contained regional populations that varied in premolar root number (22).
- 75. B. Senut, M. Pickford, C. R. Palevol. 3, 265 (2004). [CrossRef]
- 76. H. Gee, Deep Time: Cladistics, the Revolution in Evolution (Free Press, London, 1999).
- 77. C. V. Ward, A. C. Walker, M. G. Leakey, Evol. Anthropol. 7, 197 (1999). [CrossRef] [Web of Science]
- 78. We use genera to express both phyletic proximity and circumscribed adaptive systems, with ecobehavioral and morphological conditions being integral parts of the latter. This use employs the broadly defined genus Australopithecus, without recognizing the now commonly used Paranthropus (82). This is because both "robust" and "nonrobust" Australopithecus species are characterized by a commonly derived heavy masticatory apparatus (albeit to differing degrees), and also because we cannot—even to this day—be certain that the "robust" species are monophyletic.
- 79. C. O. Lovejoy, Gait Posture 21, 95 (2005). [Web of Science] [Medline]
- 80. C. O. Lovejoy, Gait Posture 21, 13 (2005).
- 81. C. O. Lovejoy, Gait Posture 25, 325 (2007). [CrossRef] [Web of Science] [Medline]
- 82. T. D. White, in The Primate Fossil Record, W. Hartwig, Ed. (Cambridge Univ. Press, Cambridge), pp. 407–417.
- 83. For funding, we thank NSF (this material is based on work supported by grants 8210897, 9318698, 9512534, 9632389, 9729060, 9727519, 9910344, and 0321893 HOMINID-RHOI), the Institute of Geophysics and Planetary Physics of the University of California at Los Alamos National Laboratory (LANL), and the Japan Society for the Promotion of Science. D. Clark and C. Howell inspired this effort and conducted laboratory and field research. We thank the coauthors of the companion papers (22-30), with special thanks to the ARA-VP-6/500 and -7/2 excavation teams, including A. Amzaye, the Alisera Afar Clan, Lu Baka, A. Bears, D. Brill, J. M. Carretero, S. Cornero, D. DeGusta, A. Defleur, A. Dessie, G. Fule, A. Getty, H. Gilbert, E. Güleç, G. Kadir, B. Latimer, D. Pennington, A. Sevim, S. Simpson, D. Trachewsky, and S. Yoseph. G. Curtis, J. DeHeinzelin, and G. Heiken provided field geological support. D. Helgren, D. DeGusta, L. Hlusko, and H. Gilbert provided insightful suggestions and advice. We thank H. Gilbert, K. Brudvik, L. Bach, D. Paul, B. Daniels, and D. Brill for illustrations; G. Richards and A. Mleczko for imaging; the Ministry of Tourism and Culture, the Authority for Research and Conservation of the Cultural Heritage, and the National Museum of Ethiopia for permissions and facilitation; and the Afar Regional Government, the Afar people of the Middle Awash, and many other field workers who contributed directly to the research efforts and results.
Received for publication 4 May 2009. Accepted for publication 8 September 2009.
The editors suggest the following Related Resources on Science sites:
In Science Magazine
- INTRODUCTION TO SPECIAL ISSUE
- Brooks Hanson (2 October 2009)
Science 326 (5949), 60-a. [DOI: 10.1126/science.326_60a]
| Full Text » | PDF »
THIS ARTICLE HAS BEEN CITED BY OTHER ARTICLES:
- The Ardipithecus ramidus Skull and Its Implications for Hominid Origins.
- G. Suwa, B. Asfaw, R. T. Kono, D. Kubo, C. O. Lovejoy, and T. D. White
(2009)
Science 326, 68-68, 68e1-68e7
| Abstract » | Full Text » | PDF »
- Careful Climbing in the Miocene: The Forelimbs of Ardipithecus ramidus and Humans Are Primitive.
- C. O. Lovejoy, S. W. Simpson, T. D. White, B. Asfaw, and G. Suwa (2009)
Science 326, 70-70, 70e1-70e8
| Abstract » | Full Text » | PDF »
- The Pelvis and Femur of Ardipithecus ramidus: The Emergence of Upright Walking.
- C. O. Lovejoy, G. Suwa, L. Spurlock, B. Asfaw, and T. D. White (2009)
Science 326, 71-71, 71e1-71e6
| Abstract » | Full Text » | PDF »
- Combining Prehension and Propulsion: The Foot of Ardipithecus ramidus.
- C. O. Lovejoy, B. Latimer, G. Suwa, B. Asfaw, and T. D. White (2009)
Science 326, 72-72, 72e1-72e8
| Abstract » | Full Text » | PDF »
- The Great Divides: Ardipithecus ramidus Reveals the Postcrania of Our Last Common Ancestors with African Apes.
- C. O. Lovejoy, G. Suwa, S. W. Simpson, J. H. Matternes, and T. D. White
(2009)
Science 326, 73-73, 100-106
| Abstract » | Full Text » | PDF »
- Reexamining Human Origins in Light of Ardipithecus ramidus.
- C. O. Lovejoy (2009)
Science 326, 74-74, 74e1-74e8
| Abstract » | Full Text » | PDF »