����

�t�B�[���h���[�N�������ɂȂ���
�P�T�D�t�i�̑��B
�i2010�N�C2011�N�x�t�B�[���h���[�N�R�W���甲���j
�@ �w�i�ƖړI
���i�̔N�ԋ��l�ʂ́C�O��������������ȑO�i1970�N��O���j��6,500�g������Q�����C�ߔN�i2005�`2009�N�j��1,500�`2,000�g���Ő��ڂ��Ă���B���ł��t�i�ނƃ����R�ނ̌������������i�e1/6�C1/25�̋��l�ʁj�B�j�S���u�i�ƃQ���S���E�u�i�͔��i�ΌŗL��ŁC���ꌧ�̓`���H�t�i�Y�V�̍ޗ��Ƃ��ČÂ����痘�p����Ă����d�v�Ȑ��Y�����ł���B���ꌧ�͂������܂ލݗ���̎�c���Y�E�������Ƃɓw�߂Ă���B
�������C�]���̃R���N���[�g�r��p�����l�H�I�Ȏ�c���Y�����͑���̘J�͂Ɣ�p��v���C�\���ʂ̎�c�Y�ł��Ȃ��Ƃ�����_���������B�����ŋߔN�́C���c�̎������������Y���𗘗p���āC�t�i�ނȂǂ̎e���𐅓c�ɕ��{���C���������܂ŗ{��������C���i�֗���������悤�Ȏ��g�݂��s���Ă���B���c�́C�e�t���̉a�ƂȂ�v�����N�g�����L�x�Ő������������߁C�t�e���̐����͗ǍD�ł���B�܂��C�O���������Ȃ����߁C�t�i�̎Y���E��������̏�Ƃ��čD�K�Ƃ���Ă���B����Ɏ��ꌧ�ɂ́C�t�����琬�ł��鐅�c�ʐς��c��ɂ���C����̃t�i���B�̖��i�Ɋ��҂�������B
 �@�@�@
�@�@�@
�������ߔN�̋��l���v���������C�ݗ����̎����ʂɖ����̒���͌����Ȃ��B�����ʂ����Ȃ������̈�ɁC���c���痬�������i���������j�t�i�t�����C�����n�_���ӂɑ�����������O�����i�I�I�N�`�o�X�C�u���[�M���j�́u�W���I�ȐH�Q�v���Ă���\�����l������B���Ȃ킿�C���i�̃t�i�ނ̑��B���Ƃ��C�������ĊO�����B���鎖�ƂƂȂ��Ă���\�����ے�ł��Ȃ��B�]���āC���c�ň琬�����t�i�t���̗�����̕��U���Ԃ���ъO�����ɂ��H�Q�̒��x�ׂ邱�Ƃ́C��c�����i�����j�̌��ʂ��������ŕK�v�ł���C��������Ȃ��Ŏ�c�̂ݕ����������邱�Ƃ́C�O�����̑��B�ɂ��Ȃ��閳�ӔC�Ȏ��Ƃƍl������B��c�̐��Y�E��������сC����ɂ���đ������O�����̋쏜�̑o���ŁC�ŋ�������i�Q��j����Ă��܂�����ł���B
�����ʼn�X�͔��i�Ή��݂̐��c�ň琬�����j�S���u�i�ƃQ���S���E�u�i�̒t�����C���c������i�ւƒʂ��鏬�͐�֗��������C�����n�_���ӂł̍̕ߔ����̌o���ω��������B�܂��C�����I�ɐ������Ă���O�������̕߂��C�����̃t�i�ނɑ���H�Q�̒��x�ׂ邱�ƂŁC�O�����̃t�i�t���ɑ���e���𐄑������i�H�Q�����Ɋւ��Ă͎����Q�Ɓj�B
�A �������@
5��21���Ƀt�i�̛z���e���i�юe�C�z����2����j�𐅓c�ɕ��{�����B���e�ɐ旧���āC���ᗑ�܂��͛z���e���̒i�K�Ŏ���ALC���F�i�A���U�����R���v���N�\���j�ɂ��W�����{�����B���{���x��10�`100��/��2�Ƃ����B���e����1�����Ԑ��c�ň琬�������ƁC6��20�`22���ɐ��c�̒��������ɐ��c�킫�̔r���H�֗����������B�Ȃ��C���c�ł̋��a�C�{�쓙�͈�؍s��Ȃ������B�������̈ꕔ���������Ɏ����A��C�̒��C�̏d�C�̍��C�얞�x�Ȃǂ��v���E�v�Z�����B
�t�i�t�����C�����O�i�\�������j����9��10���܂ŁC�����n�_���ӂŒ���I�ɍĕ߂��邱�ƂŁC�������̕��U���Ԃׂ��B�ĕߒ����́C�l��Ԃ�p���ĒP�ʎ��Ԃ�����ɍĕ߂��ꂽ�����t�������iCPUE�j����C���������ƁC�����n�_���ƂɎc�������Z�o�����B�����t���̎픻�ʁi�j�S���u�i�C�Q���S���E�u�i�j��ALC�W���a��PCR-RFLP�@�iDNA�̉���z��Ɋ�Â����@�j�ŁC�V�R���Ɛ��c�������̎��ʂ�ALC�W���̗L���ōs�����B


�ʐ^�F�t�i�̎Y���r�ɐl�H�Y���������C�e�������e����i���j�C�z���e���̌v���i�E�j


�ʐ^�F���c�֛z���e�������e�i���j�C1������̒��������ɐ��K���痬��������i�E�j


�ʐ^�F�@���K����t���𗬉������Ă���Ƃ���i���j�G�������̃t�i�t���i�E�j


�ʐ^�F�l��Ԃɂ��̕ߒ����i���j�C�͌��t�߂ł̒������i�i�E�j
�B ���ʂƍl�@
�e���c�̃t�i�t���̐����i�̒��Ƒ̏d�j�́C���{���x�Ɉˑ����ė}�����ꂽ�B�������C���{���x�ɂ��얞�x�̍��͌����Ȃ������B�������Ɋւ��Ă͕��{���x��̃T�C�Y�Ƃ̊ԂɈ�т����X���������Ȃ������B
���c���痬�������t�i�t���͗����n�_���ӂɗ��܂炸�ɁC�����㑬�₩�ɉ����i���i�j�����֕��U���C�������3�T�ԂłقƂ�Ǎ̕߂���Ȃ��Ȃ����B�܂��C�Q���S���E�u�i�̕����j�S���u�i����������̕��U�������X���ɂ������B
�j�S���u�i�ƃQ���S���E�u�i�̎�Ȑ�����͔��i�i�{�j�ł���B���̂悤�ȋ���́C�ق��̉��݈��V�тɐ�������ݗ���ɔ�ׂāC�I�I�N�`�o�X�Əo��@����Ȃ��C���B�����̍�������ƍl������B�������Ȃ���C���i�Ɉڂ��Ă���́C�I�I�N�`�o�X�ȊO�̕ߐH�ҁi�n�X�C�r���}�X�C�L�Ȃǁj�̐H�Q���邱�Ƃ��l�����邱�Ƃ���C����͂�蒷���I�Ȑ��c���̒ǐՒ��������{����K�v������B�i�I�I�N�`�o�X�ƃu���[�M���ɂ��t�i�t���̐H�Q�Ɋւ��ẮC�����ɂ܂Ƃ߂��j
 �@
�@ �@
�@
�P�U�D�O�����̐H������
�I�I�N�`�o�X�ƃu���[�M���̏����Ǔ��e���̎픻��
�i2010�N�C2011�N�x�t�B�[���h���[�N�R�W���甲���j
�@ �w�i�ƖړI
���i�Ƃ��̏W����ɂ͊O�����i�Ƃ��ɃI�I�N�`�o�X�ƃu���[�M���j�������������Ă���C�����2��ɂ��ݗ���ւ̐H�Q����эݗ���Ƃ̋��������ƂȂ��Ă���B�ݗ���̑����͎Y���Ə�����������V�тȂǂ̌Ί݈�ōs�����߁C���̎��������I�ɐ������邱���O�����̒t�e���ւ̏W���I�ȐH�Q�����O�����B
�H�������͒ʏ�C�ݓ��e��������������@����ʓI�ł���B�������C�O�����ɕߐH���ꂽ��H�҂肷��ۂɖ��ƂȂ�̂��C�����̐i�̂̊O���`�Ԃɂ��퓯��ł���B�܂��C��̓������������m�ɕ\��Ă��Ȃ��t�e���ł́C�ߐH����̑����̏��Ȃ���Ԃł����肪����C�듯��̃��X�N�������B���̂悤�Ȗ���������邽�߁CDNA�̉���z������Ƃɔ�H�҂ށE���肷����@���L���ł���B�ݗ����̔ɐB�E���������ɓ����I�ɑ��݂���O�����̐H�����C�ݓ��e���̓��肨��ѕ���DNA���͂ɂ��c�����C�ݗ����̎����Ǘ��Ƒ��B�ɗL�p�Ȓm���邱�Ƃ�ڎw���B





���@
�A �ݓ��e���̖ڎ�����
�݂̓��e����ڎ����肷��ꍇ�́C�������i�܂Ȃ��悤�ɁC�̕ߌ㒼���ɕ��o����99���G�^�m�[���𒍎˂��C�A�C�X�{�b�N�X���ɕۊǂ���i���̌�3���Ԉȓ���-20���̗Ⓚ�ɂɈڂ��ĉ�U����܂ŕۑ�����j�B�z���}������DNA�̕����𑣐i����̂ŁC�����Ǔ��e���i���Ȃǁj��DNA���͂ɋ�����ꍇ�́C�g�p���Ȃ��ق����悢�B�܂��C�O�����̍̏W�ƕ��s���Ďl��ԂⓊ�Ԃɂ��ݗ����̍̏W���s���C��������̋��ޑ��i�������j�ׂ邱�ƂŁC�O�����̐ۉa�I�𐫂��v�Z�ł���B
�ݓ��e���̎픻�ʂ́C����Ǝ��̌������ɂ���čs���B�O�`�̎c���Ă���ݓ��e���ɂ��ẮC�̒���Ғł̒����𑪒�E�L�^����B�������������O�`�ɂ��픻�ʂ�����ȏꍇ�́C������͈͂œ������������߂�B�܂��C�K�v�ɉ����Ďʐ^�Ɏc���B�܂��C�t�e���̏ꍇ�C���̌`���̔��ʂ��\�ȏꍇ������B�������i�݁i�W�{�̑������r�������j�ڎ��Ŏ킪������Ȃ��ꍇ�́u��s���v�ƋL�^���C�B���ȓ���͔�����B

�ʐ^�F���^�̃A���i2���C���j��ߐH���Ă����I�I�N�`�o�X�i���j�ƁC�X�W�G�r�i�~�������j�𑽐��ߐH���Ă����I�I�N�`�o�X�i�E�j

�B ���̍̎�ƕ���DNA�̒��o
�@�ݓ��e���ł͂Ȃ��C����DNA�͂��闝�R�́C�ݓ��e���̏ꍇ�C�ߐH����̉a�����Ƀo�C�A�X���|����߂���̂ɑ��C���̏ꍇ�͂�����x�ψꉻ����Ă��邱�Ƃɂ��B��傩��2�`3 cm�̋�Ԃ���U���Ő؊J���C���ǂ̔S�t�����������Ȃ��悤���ӂ��Ȃ���C���������s���Z�b�g�œE�o���C1.5m�k�e�}�C�N���`���[�u�ɓ���ĕ��͂���܂�-20���ŕۑ����Ă����B��U�̂��ƂɃs���Z�b�g���U�������\����C�T���v���Ԃ̃N���X�R���^�~�i���ʍ����j��h���B�������DNA���o�́C0.2 mL�ePCR�`���[�u�ɕ�2�`3 mg��DNA���o�t�iPrepMan Ultra, ABI�Ёj20��L�����ċψ�ɍ����C�T�[�}���T�C�N���[�ʼn��M����i95���C15���ԁj�B�����܂ŗ�p��C�����UṂ����S�����i3000�~g�C1���ԁj�����㐟�݂�80�{��߂���PCR�e���v���[�g�Ƃ���B�O�����̕��Ɏc������a�����R����DNA���C�ȉ��ɏq�ׂ�qPCR-SSP�@�iquantitative PCR with species-specific primers�C��ʁj����уN���[�����C�u�����[�@�iClone library�C �萫�C����ʁj�ŕ��͂���B�O�����̕��Ɋ܂܂��O�������g��DNA�i�z�X�gDNA�j�����Ȃ��悤�Ƀv���C�}�[��v���邩�CPCR�����Y�����O�������ٓI�z���F�����鐧���y�f�ŏ�������B


�C qPCR-SSP�@�ɂ��O��������DNA�̒��
0.2mL-PCR�`���[�u�ɔ������t11.5 ��L��PCR�e���v���[�g1.0 ��L�������y���UṂ��CSYBR Green I��p�����C���^�[�J���[�^�[�@�ɂ��qPCR�������s���B�{���͂ɐ旧���C���m�g���̃e�X�g�T���v����p���ėl�X�Ȕ��������Ŏ������͂��s���C���������̍œK����}��B�v���C�}�[�́C���m�z��̃}���`�v���A���C�����g�����ƂɃv���C�}�[�v�\�t�g�iPrimer3�j�ɂ��v�������ށi�����ނ��܂ށj�C�ߑ������i�b�k�ނƍ����ށj�C��̓����i�L�ށj�̂��ꂼ��ɓ��ٓI�ȃ��j�o�[�T���v���C�}�[��p����B
�܂��C�O�����ɂ��t�i�t���ւ̐H�Q�̒��x�ׂ邽�߁i�O���Q�Ɓj�C�t�i�ޓ��ٓI�v���C�}�[���g�p����B����ɁC�I�I�N�`�o�X���ٓI�v���C�}�[����уu���[�M�����ٓI�v���C�}�[��p���āC���ꂼ��u���[�M���̕�����уI�I�N�`�o�X�̕�����PCR���C�O�����Ԃ̕ߐH/��H�W�ׂ�Ƌ��ɁC�G�r�ނɂ��O�����i���j�̕ߐH�ɂ��Ă����ׂ�B
������̏ꍇ���C��������z�ʂ́C�~�g�R���h���A�c�m�`�imtDNA�j�̎���ψق̏��Ȃ����ʁi16S rRNA�j�Ȃǂ��]�܂����BmtDNA�̓Q�m��DNA��1000�{���x�̗ʂ�����̂ŁC������ł��a�����R����DNA��e�ՂɌ��o���邱�Ƃ��ł���B���o�����グ�邽�߁C�����Y���͂ł��邾���Z���ق����]�܂����BqPCR�œ���ꂽCt�l�i�����Ȑ���Threshold Cycle�j�͑��Βl�iCt = 20��1,000,000�j�ɕϊ����Č�̉�͂ɋ�����B
�D �N���[�����C�u�����[�@�ɂ��O��������DNA�̓���
�@��q��DNA���o�t���N���[�����C�u�����[�p�i�픻�ʗp�j�̃��j�o�[�T���v���C�}�[��p����PCR�������s���B��L��qPCR���l�CmtDNA������B��̔��ʂ�����s����CPCR�����Y���́C������x�̒������K�v�ł���C��ԕψق������̈悪�]�܂����B�v���C�}�[�v�̓s����CPCR�Y���ifishes�j���O�������g�i�z�X�g�j�̔z����܂ޏꍇ�́C������������邽�ߊO�������ٓI�����y�f�ŏ�������K�v������B
���͂ɐ旧���C���m�g���̎�����p���āC�v���C�}�[�̓���`�F�b�N�Ɣ��������̍œK�����s���BPCR�����ߑ��ɂ��L�����z���heteroduplex�̔�����}���邽�߁CqPCR�ŃT�C�N�����̓K�������s���B
PCR�����t��2���A�K���[�X�Q���œd�C�j�����C�ړI�o���h�̐؏o���C�����C�Z�k�C�E�����o�āCPCR�Y��5�f���[�̃����_����3�f���[�̕��������s���B����Ƀ��C�Q�[�V���������t��������PCR�Y�����R���J�e�}�[�����C�v���X�~�h��������16���ň�Ӕ���������B���C�Q�[�V������̃v���X�~�h���R���s�e���g�Z���Ɍ`���]�����CLB���V�|�n�Ŕ|�{��C�E���X�N���[�j���O�ƃR���j�[PCR�ɂ��C�����R���J�e�}�[���܂ރR���j�[��LB�|�n�Ŕ|�{���C�v���X�~�h�𒊏o������V�[�P���X��͂���B
����ꂽ�z��f�[�^����ȉ��̕��@�Ŏ�肷��B�܂��C���ނɊւ��ẮC�\�ߒ��ׂĂ��������i�ݗ����DNA�z��Ƃ̃}���`�v���A���C�����g�ɂ��ƍ�����B����ȊO�̎�ɂ����Ă�NCBI-Blast�ŋߎ��z�����������B�Ȃ��C�{�@�̓��^�Q�m���I��͂ł��邩��CDNA�z�����Ă��C���̔z��̎����傪����ł��Ȃ����Ƃ������B���̏ꍇ�͕��ތQ�i���C�ȂȂǁj�̃��x���Ŕ��ʂ���B
�E �ۉa�I�𐫁iE�j�̌v�Z
�i�I�I�N�`�o�X�̏ꍇ�j�ݓ��e������ѕ�DNA���猟�o����������̓�����C��������̋��ޑ��i�W���̒�9cm�ȉ��̌̂Ɍ���j�Ɣ�r���C�I�I�N�`�o�X�̊e�a�����ɑ���ۉa�I�𐫁iE, Ivlev�fs selectivity index�j�������ɂ��v�Z����B
E = (ri-pi)/(ri+pi)�C�������C
ri�� ������i�̐ۉa�ʒ��̑��Ηʁi���j
pi�� ������i�̊��̉a�������̒��̑��Ηʁi���j
�F �a���d�v�x�w���iIRI�j�̌v�Z
�ߐH�҂̈ݓ��e���ׁC�e�a������̌̐�����яd�ʂ���a���d�v�x�w���iIRI�CIndex of relative importance�j�������ɂ��v�Z����B
IRI��(Ni%+Wi%)*Fi%�C�������C
Ni%�� �a����i �̌̐��^�S�a�����̍��v�̐�*100
Wi%�� �a����i �̑��d�ʁ^�S�a�����̑��d��*100
Fi%�� �a����i ���o�������݂̐��^���������݂̐��i��݂������j*100
����
�����n�_�̋��ޑ��i2010�N�t�B�[���h���[�N�R���ʁj

����͎l��Ԃō̕߁i���͑S�ē��Ԃō̕߁j
�������Ԃ�6/17-9/10�C2010.
���ވȊO�ł́C�X�W�G�r�C�k�}�G�r�C�U���K�j�C�J�G���c���C�^�j�V�ނ������̏W���ꂽ�B

�ڎ�����ɂ��O�����ݓ��e������
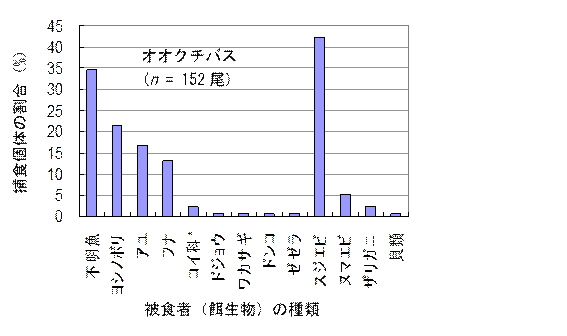
�i2010�N�t�B�[���h���[�N�R�@�������ԁF6/17�`9/10�j�\�\�I�I�N�`�o�X�̌��̐�152���̂����C��̂�22���i��ݗ�14���j�G �u���[�M��74���̑ł��C��̂�1���i��ݗ�1���j�B�I�I�N�`�o�X�̔�H�҂̂����C�R�C�Ȃ̓t�i�ȊO�̃R�C�ȋ��ނ������B�u���[�M���̔�H�҂̂����C�~�W���R�ނ�Daphnia����́C�܂��C���ނ̓��V�m�{���̂݁B���������͗c���ށC�_���S���V�C�b���C�J�����V�C�n�G�ށB���������̓��X���J�C�c���ށC���S�ށC�A�����{�ȂǁB
�i2011�N�t�B�[���h���[�N�R�@�������ԁF6/26�`8/23�j�\�\�̕ߔ����́C�I�I�N�`�o�X173���i��28���͒t���j�C�u���[�M��202���i��77���͒t���j�B�I�I�N�`�o�X�͑̒�15�`19�p�C�u���[�M���͑̒�14�p�ȉ��̌̂������̕߂��ꂽ�B�I�I�N�`�o�X�̓t�i������̒������Ԓ��C�t�i�t����26���ߐH���Ă���C���̂���ALC�W���̂���t�i��2���ł������B�I�I�N�`�o�X�̈ݓ��e���́C�X�W�G�r�C���V�m�{���C�A���C�t�i�ށC���ށi��s���j�̏��ɑ����m�F���ꂽ�B
����C�u���[�M���̈ݓ��e���͑��ށC���L�C���X���J�c�����D��ŁC�����E���������ށC�G�r�ށC�����ȂǑ��l�ȉa���������m�F���ꂽ�B�܂��C����2����Ƀt�i�t����4���ߐH���Ă���u���[�M����1�̌���ꂽ���C���̑��̌̂̓t�i��ߐH���Ă��Ȃ������B�Ȃ��C����4����ALC�W���̂���t�i�i�j�S���u�i�R�C�Q���S���E�u�i�P�j�ł������B
 �@�@�@�@
�@�@�@�@
�I�I�N�`�o�X�̑̒��Ɣ�H����̑̒��̑��֊W
���PCR�̌��ʁi2011�N�t�B�[���h���[�N�R�W���j
������t�i��DNA�����o���ꂽ�O�����i�t���͊܂܂��j�́C�I�I�N�`�o�X��130����75���i58���j�C�u���[�M����125����34���i27���j�ł������i�}�X�j�B���o�ʂ̓I�I�N�`�o�X�C�u���[�M���Ƃ��ɗ�������ɑ����X���ɂ��������C�O�҂ł͒������Ԃ�ʂ��āi8�����{�܂Łj���o���ꂽ�B�܂��C�I�I�N�`�o�X�̕����猟�o���ꂽ�t�i��DNA�ʂ́C�u���[�M���̖�100�{�ł������B����C�O�����̒t���̕�������t�i��DNA�����o���ꂽ���C���̒�ʒl�͐����ɔ�ׂĂ͂邩�ɏ��Ȃ��ʂł������B�������C�u���[�M���t���̕�����́C�I�I�N�`�o�X�t���̕�������������̃t�iDNA�����o���ꂽ�B
������ɍ̕߂��ꂽ�I�I�N�`�o�X�i�t���������j�̕���DNA�̂����C�ߑ�������DNA��72���C����DNA��22�����߂Ă����i�}10�j�B�I�I�N�`�o�X�̕������u���[�M����DNA���͂��Ɍ��o���ꂽ�B�I�I�N�`�o�X�t���ł͐ߑ�������DNA�������C����DNA�͖w�nj��o����Ȃ������B����C�u���[�M���i�t���������j�̕���DNA�̂����C�ߑ�������DNA��96���C��̓���DNA��3�����߂Ă���C���ނ�DNA�͖w�nj��o����Ȃ������i�Ȃ��C�A���a���R����DNA�͒�ʂ��Ȃ������j�B�u���[�M���t���̕������I�I�N�`�o�X��DNA���͂��Ɍ��o���ꂽ�B

�l�@
������̃t�i�t���́C�u���[�M���ɂ��H�Q�͂قƂ�ǖ����C18cm�O��̒��^�̃I�I�N�`�o�X�ɂ��ߐH�������F�߂�ꂽ�B�܂��C�I�I�N�`�o�X��30mm�ȏ�̂��Ȃ萬�������t�i�t�����ߐH���Ă���C���c�ł̗{�����Ԃ̉������邢�͕��{���x�̗}���ɂ���ė������̑̃T�C�Y��傫�����邱�Ƃ͂��܂�L���łȂ��ƍl������B �������O�ɗ����n�_���ӂŒ��^�̃I�I�N�`�o�X�̏W���I�쏜���s�����Ƃ��C�O�����ɂ�闬�����̐H�Q��ጸ�����Ō��ʓI�ł���ƍl������B
���c���痬�������t�i�t���́C�������ォ����i�֍~�͂��C������2�T�Ԓ��x�ŗ����n�_���ӂ̐��悩�炢�Ȃ��Ȃ�B�܂��C�j�S���u�i�����Q���S���E�u�i�̂ق������₩�Ɏp�������B�t�i�t���̕ߐH�̓I�I�N�`�o�X�ɂ����̂ł���C��������̕ߐH�������B�u���[�M���͖w�ǃt�i�i����ё��̋��ށj��ߐH���Ȃ��B
�ȏ�̂��Ƃ���C���c���痬�������t�i�t���̐��c�������߂邽�߂ɂ́C�����O�Ɏ��Ӑ���̃I�I�N�`�o�X���쏜����C�j�S���u�i�����Q���S���E�u�i�𑽂�����������C����Ƀt�i�ȊO�̉a�������i�G�r�ށC���V�m�{���C�A���Ȃǁj�𑝂₷�悤�Ȏ��g�݂��L���ƍl������B

�}�X�D�I�I�N�`�o�X�ƃu���[�M���̕����猟�o���ꂽ�t�i��DNA�̑��Η�

�I�I�N�`�o�X�̐ۉa�I��
|
��H���� |
���ޑ�*1 |
|
�ݒ�*2 |
E |
|
����*3 |
E |
|
�A�� |
3 |
|
28 |
0.98 |
|
6*4 |
0.95 |
|
���V�m�{���� |
78 |
|
54 |
0.71 |
|
28 |
0.77 |
|
�t�i�ށC�t�� |
770 |
|
33 |
-0.46 |
|
9 |
-0.60 |
|
�h�W���E |
13 |
|
1 |
-0.21 |
|
|
|
|
���J�T�M |
1 |
|
5 |
0.95 |
|
|
|
|
�I�I�N�`�o�X�C�t�� |
40 |
|
0 |
-1.00 |
|
|
|
|
�u���[�M���C�t�� |
50 |
|
0 |
-1.00 |
|
|
|
�ۉa�I���w�� (E) �̌v�Z�̓C�u���t�̕��@�ɂ��B
�T���v���T�C�Y (ri+pi) ��6 �����̎�͂d���Z�o�����B
���̃T�C�Y�̑傫���� (�W���̒�9 cm �������) �͌v�Z���珜�O�B
*1 ��������ō̏W���ꂽ�̐�
*2 �I�I�N�`�o�X�̈ݓ�����E�o���ꂽ�̐��B
*3 �I�I�N�`�o�X�̕�������P�����ꂽ�N���[�����B �I�I�N�`�o�X�ƃu���[�M���̔z��͐����y�f�����ŏ��O�������߁C�N���[���Ɋ܂܂�Ă��Ȃ��B
*4 �A���ƃ��J�T�M�̍��v�B

�I�I�N�`�o�X�E�u���[�M���̐H���̂܂Ƃ�
�I�I�N�`�o�X�̐H��
l ���ށi�A���C���V�m�{���C�Ȃǁj�ƃG�r�ށi�X�W�G�r�C�e�i�K�G�r�j�𑽂��ߐH�B
l �L�ނ�A���͖w�ǕߐH���Ȃ��B
l �傫���a������ߐH����X��������i�t���ނ͕ߐH���Ȃ��j�B
l �a�����ɑ���I�𐫂́C�ߐH�҂̚n�D���ƖO�H�x�C�a�����̐H���C�s���l���C�߂��Ղ��C�������Ղ��C�傫�����W����B
�u���[�M���̐H��
l �A���a���𑽂��ߐH�B
l ���������C�L�ށC�~�W���R��Daphnia spp.�C���������C�k�}�G�r�C�ȂǑ��l�Ȑ�����ߐH�B
l ���ނ͂قƂ�ǕߐH���Ȃ����C�����͍D��ŕߐH�B
l �����ȉa�����i�~�W���R���̃v�����N�g���j�������ߐH�B
�P�V�D�t�B�[���h���[�N�̌��ʂ��l�@����ɓ�������
�I�I�N�`�o�X�̉a�����ɑ���ۉa�I�𐫂�����ƁC�t�i�t���ɑ���I�𐫂����Ȃ�Ⴂ���Ƃ�������B����́C�t�i�̑̃T�C�Y�̉e���C���Ȃ킿�I�I�N�`�o�X����^�̉a�����ɑ��āC�������̑I�𐫂����邱�Ƃ�����ƍl�����邪�C���������^�̃��V�m�{���ނ��C�t�i�t���Ƃ͑ΏƓI�ɑ����ߐH����Ă��邱�Ƃ���C�傫���ȊO�ɂ��I�I�N�`�o�X���t�i�t������������������̂�������Ȃ��B
�t�i�t���Ɠ����悤�ȑ̌`���������ɁC�^�i�S�ނ�����B�^�i�S�ނ͉p����bitterlings�i�ꂢ�����̈Ӂj�ƌĂ��悤�ɁC�ꂢ���������I���B���{�ł��^�i�S�ނ��j�K�u�i�i�ꕩ�j�ƌĂԁB����́C�^�i�S�ނ��ߐH�҂̐H�Q���瓦��邽�߂̎�i�̂ЂƂƂ����Ă���B�I�I�N�`�o�X�Ȃǂ̕ߐH�҂��C�^�i�S�ނƃt�i�t������ʂł��Ȃ��Ƃ���C�t�i�t���̕ߐH�����Ȃ����R�̂ЂƂ��^�i�S�ނւ̋[�ԁi�H�j�Ƃ����\�����l������B���������i�Ń^�i�S�ނ̎����ʂ��������Ă��鍡���C�I�I�N�`�o�X���^�i�S�ނ̋ꂳ���o������w��ł���Ƃ͍l����B�ނ���C�^�i�S�^�̑̌`���������ނ�{�\�I�ɔ�����A�v���I���I�Ȑۉa�K���������Ă���̂�������Ȃ��B
�������ȏ�q�ׂ����Ƃ͑z���ł���C�����ɉ߂��Ȃ��B�Ȋw�I�ɕ������l���邱�ƂŁC���̂悤�ȉ��������X�ɐ��܂��B�����āC���̂悤�ȉ����͎����ɂ���Č����邱�Ƃ��ł���B���ꂪ�C�����̂͂��܂�ł���B�t�B�[���h���[�N�Ńf�[�^�����W������C���̃f�[�^�����Ƃɂ��đ��l�ɑz�����߂��点�邱�Ƃ���ł���B�f�[�^�������I�Ɏ��W���C�������ĕ��邾���ł́C���ɂ�����Ȃ��t�B�[���h���[�N�ɂȂ��Ă��܂��B���E�ɂP�����Ȃ����������������Ă���M�d�ȁi�閧�́j�f�[�^�����ƂɁC���낢��Ƒz�����C�V�����ɂȂ���悤�ȉ����𗧂Ă邱�Ƃ��C�����̊y���݂ł���C������ł���B
�����ő�Ȃ��Ƃ́C���}�Ȕ��z�ł͂Ȃ��C��Ȕ��z��t�]�̔��z���ł��邩�C�Ƃ����_�ł���B����́C�o�J�Ȕ��z�Ǝ���d�̂��Ƃ��������C�Ƒn���ɂȂ���d�v�Ȉ���ł���B�펯�⑼�l�̖ڂɂƂ���邱�ƂȂ��C�I���W�i���ɑ�_�ɔ��z���邱�Ƃ��d�v�ł���B���̂��߂ɂ́C�܂������ł�������ƍl���C�Y�ނ��ƁB���悭��ƔY�ނ��ƁC�v���߂邱�Ƃ́C�����ɂ����đ�Ȕ\��/�˔\�ł���B
���ɏd�v�Ȃ̂��C�搶���͂��߁C�����̐l�Ƌc�_���邱�Ƃł���B�Ȃ��c�_���d�v�Ȃ̂��B����́C�l�X�Ȃ��̂̌����C�l������m��i�C�t���j���߂ł���B�Ƃ�悪��̍l���́C��͂������ア�i�Ƃ��ɎႢ�l�j�B��s�������͂��ߑ����̂��Ƃɂ��āC�搶���y�͗ǂ��m���Ă���i���ʁC�m���ʂɂ����āC��҂̔�ł͂Ȃ��j�B�����Ɋւ��ẮC���łɕ������Ă��邱�Ƃ��ꐶ�����Ɍ������Ă��d�����Ȃ��i�قƂ�lj��l���Ȃ��j����ł���B
�������鉿�l�̂���u�悢�����v�́C��l�̂Ђ�߂���a�V���C���邢�͎v�����݂����łȂ��C�c�_�̒��Ő�������邱�Ƃ��d�v�ł���B
�P�W�D���w�W��`�q�Ɗ��f�f
�i2007�N�x�t�B�[���h���[�N�R�W�����p�j
���R����̊��]���́C�����ɐ������鐅�������C���Ғœ����C���ނȂǂ̊��w�W���������Ƃɍs���邱�Ƃ������B�������Ɋ�Â����̕��@�́C���ʂȒm���C�Z�p�C���͋@��Ȃǂ�v���Ȃ����Ƃ�̏W�̊y��������C������ΏۂƂ������R�ώ@��Ȃǂł���ʂɍs���Ă���B���̕��@�́C�������鐶���̎�ނƌ̐��i����ѕ⑫�w�W�Ƃ��āC�傫����N��\���Ȃǁj���w�W�Ƃ��Ă���C���ԓI��ԓI�ϓ��������ʂ��o�����Ƃ��ł���B�������C�̐��Ɋ�Â����̂悤�Ȏw�W�͐����ȊO�̊��v���C�Ⴆ�C��ԊW�C�a�����̋����C�V�G�C�G�߁C�����C�����C�ꎿ�C�ɐB�ꏊ�Ȃǂɂ��傫���e�������B
����C����̊������x��f�f������@�Ƃ��āC�ߔN�͖��Ғœ����C���������C�v�����N�g���C���ނȂǐ��������̑̓��̎w�W��`�q�i�}�[�J�[��`�q�j�̔����x�ׂ���@����������Ă���B��������X�g���X�ɕq���ɉ�������w�W��`�q�̔����x�ׂ邱�ƂŁC���w�����C�Ⴆ�C��𐫗L�@�������C�_��C���z�������C�d�����Ȃǂɂ�鐅�托�����C��r�I�����̒i�K�Őf�f����̂��ړI�ł���B���̂��߂̎w�W��`�q�̌����Ɣ����x�̉�͂́C���f�f�̗L�p�Ȍ����Z�@�Ƃ��Ċ��҂���Ă���C�}�C�N���A���C����p���ĉ����������ƂɃ}�[�J�[��`�q�̓����i�߂Ă���B���̂悤�Ȋ��Ő������i���������j���̓��̈�`�q�����ɗ^����e���ׂ�g�L�V�R�Q�m�~�N�X�̕���́C�������w�̏d�v�Ȍ����̈�ƂȂ��Ă���B
����̃t�B�[���h���[�N�ł́C������������Ă���X�g���X�x���w�W��`�q�̔����x�����Ƃɐf�f���邱�Ƃ����݂��B�w�W��`�q�Ƃ��đI�������̂́C��Ōn�̐��_���y�f�ł���CYP1A1�ŁC�g�p���������͌����i���̃t�B�[���h���[�N�̃t�B�[���h�̈�j�ő����̏W����郈�V�m�{���ƃX�W�G�r�ł���B��`�q�̔�����͂̓��A���^�C��PCR�Œ��ׁC�x�[�^�A�N�`���Δ�ʂƂ��ċ��߂��B�Ȃ��C���ׂ�CYP1A1�̉���z��͖��m�ł������̂ŁC�܂����̉���z��ׂ�Ƃ��납�璅�肵���B
�����̗���͈ȉ��̒ʂ�F�@�c�m�`�f�[�^�[�x�[�X�iNCBI�CEnsembl�j�ɂ��߉���̉���z��̌���➔ �}���`�v���A���C�����g➔ �k�d�v���C�}�[�쐬➔ RNA�̒��o�C�����C�Z�x����➔ RT-PCR�C�d�C�j���C�N���[�j���O➔ �X�N���[�j���O�C�咰�۔|�{➔ �v���X�~�h���o�C����➔ �V�[�P���X➔ ���A���^�C���p�v���C�}�[�쐬➔ ���A���^�C��PCR➔ �f�[�^��́B�Ȃ��C���̎����͈ȉ��ɏq�ׂ�_��\�I�����Ƃ��킹�Ď��{�����B
�_�������ƂƂ��ɐ���ɗ�������ƁC�����ɐ�������l�X�Ȑ������Ő������ɖ\�I�����B���̔Z�x���Ⴂ�ꍇ�́C��Ōn�̈�`�q���������債�C���̂��̓��ɐN������ŕ��̉�Ŕr���̂��߂̋@�\�傷��B�ŕ��̗����ʂ������ꍇ�́C�����͂��̐��悩�狏�Ȃ��Ȃ�i������j���C�������͎��S����B
�����2��ނ̎s�̔_��i�_��`�C�_��a�Ƃ���j��p���Ď������ł̖\�I�������s���C�ˎ�������ш�`�q�����ׂ��B�܂��C24���Ԍ�̝ˎ��������ƂɓŐ��ׁC�����c�����̂ɂ����Ă͎w�W��`�q�iCYP�j�̔����ׂ邱�ƂŁC�����̊��X�g���X�x��f�f�����B�@�\�I�����ɂ̓��V�m�{���i���j�ƃX�W�G�r��p�����B
�@�����́C300ml�e�̃r�[�J�[�ɁC�Z�x��5�i�K�ɕω��������_��n�t�����C�e�r�[�J�[�Ƀ��V�m�{���܂��̓X�W�G�r��6�������e���C�キ���C���Ȃ���24���Ԗ\�I�������B
�@���ʂ�\�S�Ɏ����B���V�m�{���ł͔_��`�̔Z�x��5ppm�ȏ�̗n�t�Ő��c�����������ቺ�����B�_��a�ł�50ppm�̔Z�x�ł��ˎ��͖��������B����X�W�G�r�ł́C�_��`�C�a���ɁC0ppm���Ȃ킿�_������Ȃ������ΏƋ�ȊO�́C�S��őS�̂����S�����B�@���������āC�Z�x�������čēx�\�I�������s�����B�@�������C�Ď����ł̍Œ�Z�x�i0.03ppm�j�ɂ����Ă��S�̂����S�����B����̎����ŁC�����_��ł��C�X�W�G�r�̓��V�m�{���ɔ�ׂĂ͂邩�ɔ_��Ɏア�Ƃ������ʂ��B
�Ƃ��ɔ_��a�̎����ł̓��V�m�{����50ppm�ł��S�Đ������Ă����̂ɑ��C�X�W�G�r��0.03ppm�ł��S�Ď��S�����B�_��a��LC50�̓R�C��2.8ppm(48���ԁj�C�~�W���R��0.01ppm(3���ԁj�ƕ���Ă���C���ނ̂ق����ϐ�������悤���B����C�_��`��LC50�́C�~�W���R��0.18ppm(48����)�ƕ���Ă���̂ɑ��C����̃X�W�G�r��0.03ppm�őS�Ď��S�������Ƃ���C�X�W�G�r�̓~�W���R�������̔_��ɑ���ϐ����Ⴂ���Ƃ����@���ꂽ�B
��L�̔_��\�I�����ŁC�����c�������́i���V�m�{���̂݁j�̊̑���E�o���C�g�D���̉�Ōn�y�f�̈��iCYP1A1�j�̔����x��ΏƋ�i�_��0ppm�j�̂���Ɣ�r�����B�̑��̌Œ�ɂ�RNAlater��p�����BCYP1A�̔����́C�_��`�ɂ���Ė��炩�ɑ����������C�̍�����ł������i�}�V�j�B����C�_��a��CYP1A�̔�����U�����Ȃ����ʂƂȂ����B�X�W�G�r�́C�\�I�����őS�̂����S�����̂ŁC��`�q�̔�����͍͂s��Ȃ������B
�����ō̏W�������V�m�{���̊̑���CYP1A�̔����́C�\�I�����̑ΏƋ�i0ppm�j�̂��̂Ɣ�ׂāC���ϒl�ɂ����Ă�⍂�����C�L�Ӎ��͌����Ȃ������B������C�����̐�����CYP�̔�����U������悤�ȕ����͊܂�ł��Ȃ����Ƃ��������ꂽ�B

�ʐ^�F�_��\�I�̊ȈՎ������u
�\�S�D�_��`�Ɣ_��a�̋��Ő��ƃG�r�Ő��i24���Ԗ\�I�j
|
|
���V�m�{�� |
�X�W�G�r |
||||||||
|
�Z�x�ippm�j |
0 |
2 |
5 |
10 |
50 |
0 |
0.03 |
0.1 |
0.3 |
1 |
|
�_��` |
0 |
0 |
5 |
6 |
6 |
0 |
6 |
6 |
6 |
6 |
|
�_��a |
0 |
0 |
0 |
0 |
0 |
0 |
6 |
6 |
6 |
6 |
�e��C�S6�����̎��S�̐���\���B

�}�V�DCYP1A��`�q�̔�����́F
�_��`�i0�`50ppm�j�C�_��a�i0�`5ppm�j��24���Ԗ\�I�������V�m�{���̊̑���CYP1A��`�q��mRNA���x���B�����x�̓x�[�^�A�N�`���Δ�ʂƂ��ĕ\���B�e�J�����͕��ϒl�C�G���[�o�[�͕W�����C�����͒��ׂ��̐�(n)��\���B�@�u�����v�́C���͐�ō̏W�������V�m�{���̓���`�q�̔����x��\���B